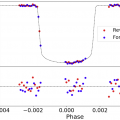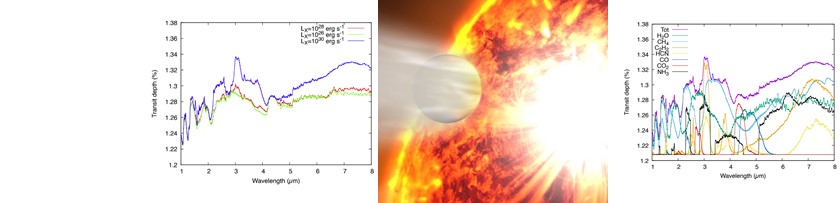
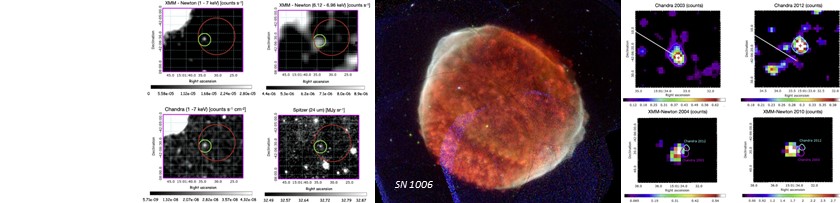

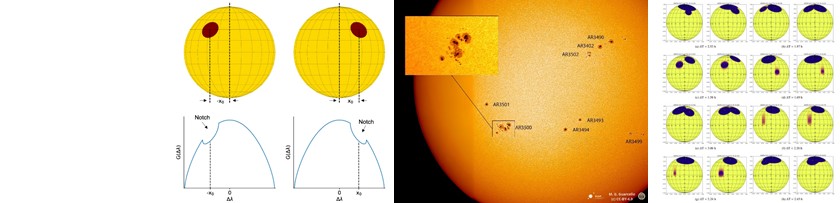




Ogni appassionato di fantascienza, in particolare della saga di Star Wars, si è almeno una volta chiesto se pianeti che orbitano attorno a due stelle, come Tatooine, siano davvero possibili. In realtà, si tratta di una domanda molto interessante anche dal punto di vista astronomico. Nella nostra Galassia, infatti, si stima che almeno un terzo delle stelle di piccola massa
» Read more